FNR-104
Infection of tree seedling roots with certain types of soil fungi can increase seedling growth as much as 400 percent. In fact, some tree species must have fungi on their root systems to develop "normally." These types of fungi are called mycorrhizal fungi and are credited with improving a seedling's ability to withstand extremely droughty soil conditions and low soil nutrient levels. In fact, hardwoods and conifers which have mycorrhizal fungi on their roof systems survive and grow under soil moisture conditions where non-mycorrhizal seedlings die. Research on several hardwood seedling species indicates that mycorrhizal seedlings are capable of growing as large as non-mycorrhizal seedlings but at one-half the level of soil nutrients.
Forest scientists are working to increase yields of fuel and fiber which are essential to human needs and which must be produced on a diminishing quantity of forest land. Largely as a result of research on forest trees, a new technology that attempts to use mycorrhizal fungi for mankind's benefit is emerging. Plant scientists have learned to think of a continuum system-the soil, the plant, and the atmosphere. This thinking should be expanded to include another component-the mycorrhizal fungi.
Mycorrhizae are specialized, root-like organs formed as a result of the symbiotic (mutually beneficial) association of certain fungi with the roots of higher plants. Specific fungi grow upon and vigorously invade portions of the root in that area of the root system that is primarily responsible for nutrient absorption. The term "mycorrhiza" literally means "fungus- root"and is used to denote these particular associations of roots and fungi.
The vast majority of economically important plants form mycorrhizae. Without mycorrhizae, most of our important tree species could not long survive in the dynamic, fiercely competitive biological communities that inhabit forest soils. Furthermore, the mycorrhizal condition is the rule, not the exception, in nature. The physical appearance or morphology of mycorrhizae varies among plant species, and each plant species tends to have characteristic groups of fungi capable of producing mycorrhizae. On the basis of their morphology, these associations are currently divided into two major groups: ectomycorrhizae and endomycorrhizae. Of the two, endomycorrhizae are by far the most common, but ectomycorrhizae are formed on some very important families of forest trees.
Ectomycorrhizae are formed by fungi belonging to the higher Basidiomycetes (mushrooms and puffballs), Ascomycetes (cup fungi and truffles, and Phycomycetes in the family Endogonaceae. The host plants of these fungi are predominantly trees such as pine, hemlock, spruce, fir, oak, birch, beech, eucalyptus, willow, and poplar. Many species of fungi may be involved in the ectomycorrhizal association of a forest, a single tree species, an individual tree seedling, or even a small segment of lateral root. As many as three species of fungi have been isolated from an individual ectomycorrhizal root cluster.
Ectomycorrhizal infection is initiated from spores (reproductive structures) or hyphae (vegetative growth structures) of the fungal symbiont which are in the soil around feeder roots. Growth of the mycorrhizal fungi on the surface of short roots is stimulated by exudates from the roots. Fungal mycelia (strands of hyphae growing together) grow over the feeder root surfaces and form an external mantle or sheath (Figure 1). Following mantle development, hyphae grow intercellularly, or between the cells, forming a network of hyphae (Hartig net) around the root cortical cells. Physical or chemical properties of roots restrict the hyphae of all mycorrhizal fungi to the cortex and meristematic (capable of actively dividing) cells of the root tip. The exact mechanism for this resistance to hyphal penetration by other parts of the plant is not known. The Hartig net, which may completely replace the tissue between cortical cells, is the major distinguishing feature of ectomycorrhizae.
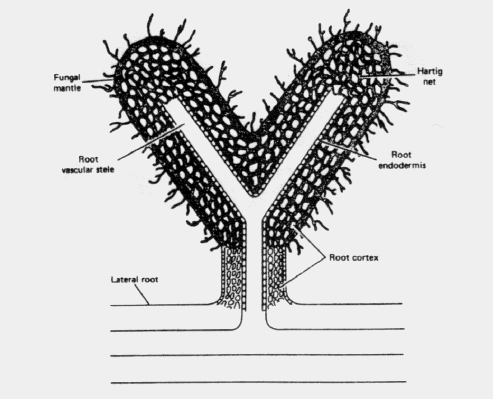
The growing, absorbing roots of the species of forest trees that become enveloped by ectomycorrhizal fungi are subject to growth regulatory substances produced by the fungi. A hormone, auxin, produced by the fungi, modifies subsequent root growth, retarding elongation of short roots and frequently initiating dichotomous branching in pine (Figure 2). Such roots are usually less than 0.5 cm in length and do not have a root cap. The cortical cells are orient- ed differently from those of non-mycorrhizal roots, and the presence of intercellular hyphae causes a swollen appearance. Ectomycorrhizae may cause other morphologically distinct root characteristics; but in all cases, the hyphae on short roots often radiate from the fungus mantle into the soil, thereby greatly increasing the absorbing potential of the roots.

The endomycorrhizal fungi are the most widespread and important root symbionts. They are found throughout the world in both agricultural and forest soils. They occur on most families of hardwood and on some conifer tree species and include, perhaps, all agronomic and horticultural crops. Among the trees with this type of mycorrhizae are maples, elms, ash, walnut, sweetgum, yellow poplar, sycamore, cottonwood, black locust, and willow. Although endomycorrhizae commonly occur, they have been ignored by many plant scientists because they have little effect on root morphology and are difficult to detect in roots (Figure 3).

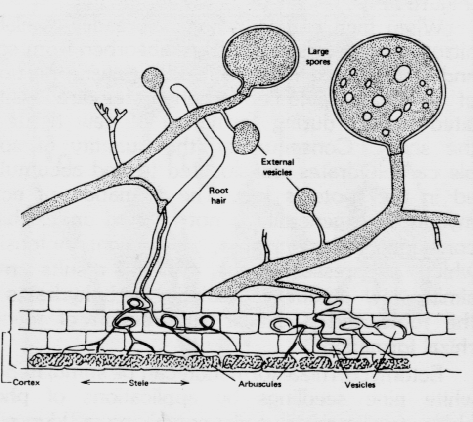
The fungi invade the cortex, but not the endodermis or stele, of the feeder roots (Figure 4). The colonization does not alter root morphology, and the sheath of fungus mycelium common to ectomycorrhizae is lacking. Under a microscope, endomycorrhizae (commonly termed Vesicular-Arbuscular) are diagnosed by the presence of vesicles (terminal, spherical structures that contained oil droplets) and arbuscules (complex structures formed by repeated dichotomous branching of hyphae) in the cortical cells of differentially stained feeder roots. Mycelia emanate from the infected root to form a loose network in the rhizosphere and adjacent soil.
The Vesicular-Arbuscular (VA) mycorrhizae are formed by certain fungal species of the family, Endogonaceae. They do not produce large, above-ground fruiting bodies or wind- disseminated spores as do most ectomycorrhizal fungi. The spores are often grouped in sporocarps or appear singly in the soil or in plant roots. These sporocarps or spores usually occur beneath the leaf litter or in the mineral soil, occasionally becoming airborne during dust storms in semi-arid regions. The fungi are more commonly spread by growing from feeder root to feeder root and, at times; are disseminated by moving water, soil, insects, and animals.
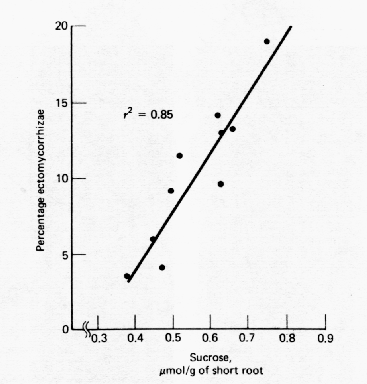
Environmental factors may influence mycorrhizal development by affecting either the tree roots or the fungal symbionts. After the formation of a receptive tree root, the main factors influencing susceptibility of the root to mycorrhizal infection appear to be photosynthetic potential and soil fertility. High light intensities and low to moderate soil fertility enhance mycorrhizal development, while the opposite conditions may reduce or even prevent mycorrhizal development. These factors may influence the biochemical status of the root by controlling the level of reducing sugars, or they may affect the synthesis of new (susceptible) feeder roots (Figure 5).
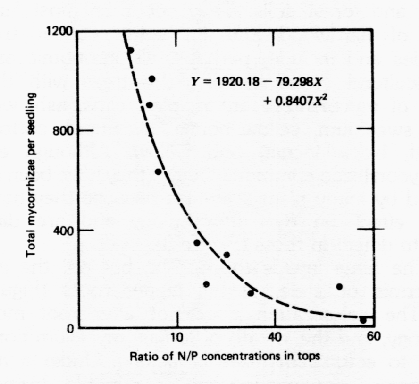
When high concentrations of readily available nitrogen and phosphorous are absorbed from soils and translocated upward to the growing portions of the plant, soluble carbohydrates are assimilated rapidly during formation of new tissue in the shoot. Consequently, the quantity of soluble carbohydrates translocated to and accumulated in the roots is low. The formation of ectomycorrhizae generally is not favored under these conditions. Furthermore, low light intensity, which suppresses shoot growth, results in a similar low amount of soluble carbohydrates in the roots and a similar suppression of mycorrhizal formation.
Ectomycorrhizal formation can be stimulated on white pine seedlings by applications of phosphate fertilizers to soils containing a low population of relatively inactive mycorrhizal fungi. It appears possible that the growth suppression often associated with nitrogen fertilization to phosphorous deficient soils results from a reduction in mycorrhizal development (and phosphorous absorption) due to the high nitrogen:phosphorous ratio in host plant tissue (Figure 6). The effects of soil fertility and fertilizer additions on the development of endomycorrhizae appear to vary with the original fertility of the soil before fertilizers are added and the nutrient content of the host plant.
There is no evidence that light has any direct effect on mycorrhizal development in soils; however, temperature can have a profound effect on the growth of certain mycorrhizae. Optimum temperatures for mycelial growth lie between 18C and 27C for the majority of fungal species. Other fungi have a wider temperature tolerance evidenced by alpine and arctic timber lines which are formed by ectomycorrhizal tree species. Such ectomycorrhizal fungi as Pisolithus tinctorius which develop at soil temperatures of 34C or higher, offer advantages in reforestation of adverse sites.
Apparently all mycorrhizal fungi require oxygen to survive, and it is presumed that mycelial growth decreases with lowered oxygen tensions; that is, as soils become flooded the levels of oxygen decline and fungal mycelial growth decreases. It is also likely that the requirements of mycorrohizal fungi for nutrient elements are not greatly different from those of the host plants, although little research has been conducted on this point. It is generally conceded that formation of ectomycorrhizae on tree roots is greatest under acid conditions.
The presence of antagonistic soil microorganisms can influence survival of the symbiont as well as root growth of the host plant. Fungicides used in plant disease control can inhibit mycorrhizal fungi under some conditions or may stimulate mycorrhizal development in others by reducing microbial competition. Fungitoxicants applied to young seedlings in nurseries may have an inhibitory effect on the development of VA mycorrhizae. Alteration of the host metabolism as a result of application of systemic fumitoxicants can suppress fungal invaders, but the effect appears to be short-lived.
The dependence of most species of forest trees on mycorrhizae to initiate and support healthy growth has been most strikingly illustrated by the problem encountered in introducing trees in areas devoid of the mycorrhizal symbionts. An example is the failure of monterey pine in western Australian nurseries that lacked mycorrhizal fungi. The seedlings grew normally only after soil from the healthy pine stand was added to the beds and ectomycorrhizae formed. The former treeless plains of the United States offer another example of an area which is deficient in ectomycorrhizal symbiotic fungi.
The observed benefit of mycorrhizae in the growth and development of trees has been ascribed to several factors. Among these factors are 1) increase in nutrient and water absorption by virtue of an increased absorptive surface area resulting from the formation of short roots and by mycelia permeating the soil in the vicinity of short roots; 2) increase in nutrient mineralization through biological weathering; and 3) increase in feeder root longevity by providing a biological deterrent to root infection by soil pathogens. There is general agreement that mycorrhizae increased the capacity of infected plants to absorb nutrients which can be especially important on infertile or adverse sites, such as mine spoils. Carbon compounds synthesized in the green tissue of the host plant not only nourish the host itself but are a source of carbon for fungal mycelia. In turn, soil-derived nutrients absorbed by the mycelia in the soil pass into host tissue. Ectomycorrhizae are able to absorb and accumulate various elements such as nitrogen, phosphorous, potassium, and calcium in the fungus mantle and then translocate these elements to host plant tissue. The importance of mycorrhizae to nutrient absorption is illustrated in Figure 7.

The importance of VA endomycorrhizae to phosphorous nutrition is illustrated in Figure 8. The VA endomycorrhizae significantly increased growth of plants on soil deficient in readily available phosphorous. The main effect of plant response to these symbionts is increased efficiency of nutrient uptake. Thus, if plants are colonized with the appropriate VA endomycorrhizal fungi, the estimate of production potential of a soil and its fertilizer requirement might change radically. However, the importance of mineral solubilization by mycorrhizal fungi to the nutrition of the host plant is not well understood. There seems to be little evidence that VA mycorrhizae can exploit less soluble forms of phosphates; rather they increase the efficiency of available phosphorous absorbed from the soil.

Mycorrhizal fungi have been reported to afford protection from attack from pathogenic fungi to delicate root tissue. This protection apparently results from the fungal mantle serving as a physical barrier to infection. Even without the mantle, root cortex cells surrounded by the Hartig net also are resistant to pathogens. In addition to a physical barrier, there is an antibiotic mechanism of resistance. This resistance is derived from a chemical substance identified as diatretyne nitrile. There is also evidence that the symbiotic fungus may supply the higher plant with more than inorganic nutrients from the soil. They may also provide the host plant with growth hormones, including auxins, cytokimins, gibberellins, and growth regulating B vitamins.
Research on mycorrhizal associations on trees growing in natural environments has revealed that 1) any ectomycorrhizae on tree seedlings are better than none, and 2) some species of ectomycorrhizal fungi under certain environmental conditions are more beneficial than others. Appropriate methods of selecting, propagating, manipulating, and managing the most desirable fungal symbionts can lead to improvement in tree survival and growth on a variety of forest sites. Practical use of ectomycorrhizal fungi can be of major significance n forest regeneration. Inoculations of planting stock with specific ectomycorrhizal fungi can increase survival and growth of seedlings planted on cutover lands, former treeless areas, and disturbed or adverse sites such as mining spoils. Most of the work on inoculation in ectomycorrhizal fungi has been done in nurseries that produce bare root or "containerized" tree seedlings. Another promising application, however, is inoculation of seed for broadcasting on sites that are too remote or too rough for convenient planting of seedlings.
Research on endomycorrhizae of trees is quite limited. Existing data is limited to trees planted on adverse sites. Researchers have found abundant endomycorrhizae on roots of a variety of herbaceous plants on coal spoils in Pennsylvania. They concluded that endomycorrhizae were, in fact, essential for the survival and growth of herbaceous plants on these coal spoils. Some researchers have found that endomycorrhizae significantly increased survival and growth of fourwing saltbush on strip-mined coal spoils in New Mexico. Endomycorrhizal seedlings were 1.5 times larger than non-mycorrhizal seedlings after the second year. Other researchers have identified endomycorrhizae on planted sycamore, sweetgum, maple, and black alder or kaolin and Fuller's earth spoils in Georgia. The endomycorrhizae on these trees were probably on the roots before planting on the spoil; it is biologically significant, however, that the endomycorrhizal fungi persisted.
Three types of materials have been used for mycorrhizal inoculation in nurseries 1) soil from natural forests or nurseries; 2) mycorrhizal seedlings; and 3) pure cultures of mycorrhizal fungi. Of the three methods, pure cultures of mycorrhizal fungi appear to hold the most promise as a method of inoculating nursery soils. Although commonly used experimentally, this technique has been rarely used in larger scale nursery operations because of the difficulties in mass producing qualify inoculum. Commercial production has recently been initiated and expansion in the use of inoculum can be expected in some areas. Pure culture techniques for mass producing inoculum of endomycorrhizal fungi are lacking. However, on a limited scale colonized soil or plant roots have successfully been used as the inoculum source. Most scientists feel that inoculation during the seedling and nursery stage offers the best possibility for promoting seedling survival and rapid early growth after outplanting. Inoculating seedlings at the time of outplanting has demonstrated no significant gains over non-inoculated seedlings. Because of the abundance of natural inoculum in a forest soil environment, inoculations with specific mycorrhizal fungi subsequent to out-planting offer no advantage to the target host plant.
Forest scientists have known about possible benefits of ectomycorrhizae for some time, but only in the last few years has the significance of endomycorrhizae begun to be appreciated. These structures are common not only in most species of forest trees, but also in most crop plants. Therefore, researchers cannot help but believe that mankind will benefit from the manipulation of these symbiotic associations.
RR 9/93
Cooperative Extension work in Agriculture and Home Economics, State of Indiana, Purdue University and U.S. Department of Agriculture cooperating: H.A. Wadsworth, Director, West Lafayette, IN. Issued in furtherance of the acts of May 8 and June 30, 1914. The Cooperative Extension Service of Purdue University is an equal opportunity/equal access institution.